

“(…) un perfetto apparato prensile rappresenta un aiuto prezioso anche per i processi più elevati dell’intelletto (…)
Herbert Spencer Principles of Psychology, 1855
Il filosofo britannico H. Spencer mise in evidenza la connessione tra l’evoluzione della mano prensile con lo sviluppo delle capacità cognitive, non solo per quanto concerne la capacità di comprensione di un oggetto ma, quindi, anche per elaborazioni comportamentali più sofisticate.
Azione e percezione
Un’azione è la risultanza, infatti, dell’interazione fra i sistemi motori e i sistemi sensitivi. In moli casi, perciò, “azione e percezione non sono separabili”[1]. È osservabile nell’esperienza di tutti il fatto che molte delle nostre azioni sono influenzate dal modo di percepire, mi riferisco non solo a processi relativamente semplici dell’afferrare un oggetto (come analizzeremo in questa lezione) ma, estendendo il concetto, al fatto di percepire una situazione, un contesto, una relazione (con noi stessi e con l’altro) e compiere delle azioni conseguentemente al proprio modo di percepirli e interpretarli, processo questo che, come sperimentato in Sigmasofia, è tutt’altro che un’azione meccanica, ma appunto determinata dal livello di autoconsapevolezza. Cercherò, quindi, di esporre parte del funzionamento neurologico del movimento estendendolo ai “processi più elevati dell’intelletto”.
Abbiamo precedentemente visto come il midollo spinale sia la struttura fondamentale di connessione tra la corteccia e il resto del corpo (in realtà un unico sistema di funzionamento), in particolare ogni movimento è possibile grazie ai motoneuroni del corno ventrale del midollo spinale, direttamente connesso al corno posteriore (area delle afferenze sensorio-percettive). Il movimento volontario, come il semplice gesto di prendere una tazzina di caffè, implica in realtà meccanismi nervosi sorprendentemente complessi che richiedono steps preparatori nella corteccia parietale e frontale, premotoria e motoria.
Aree motorie
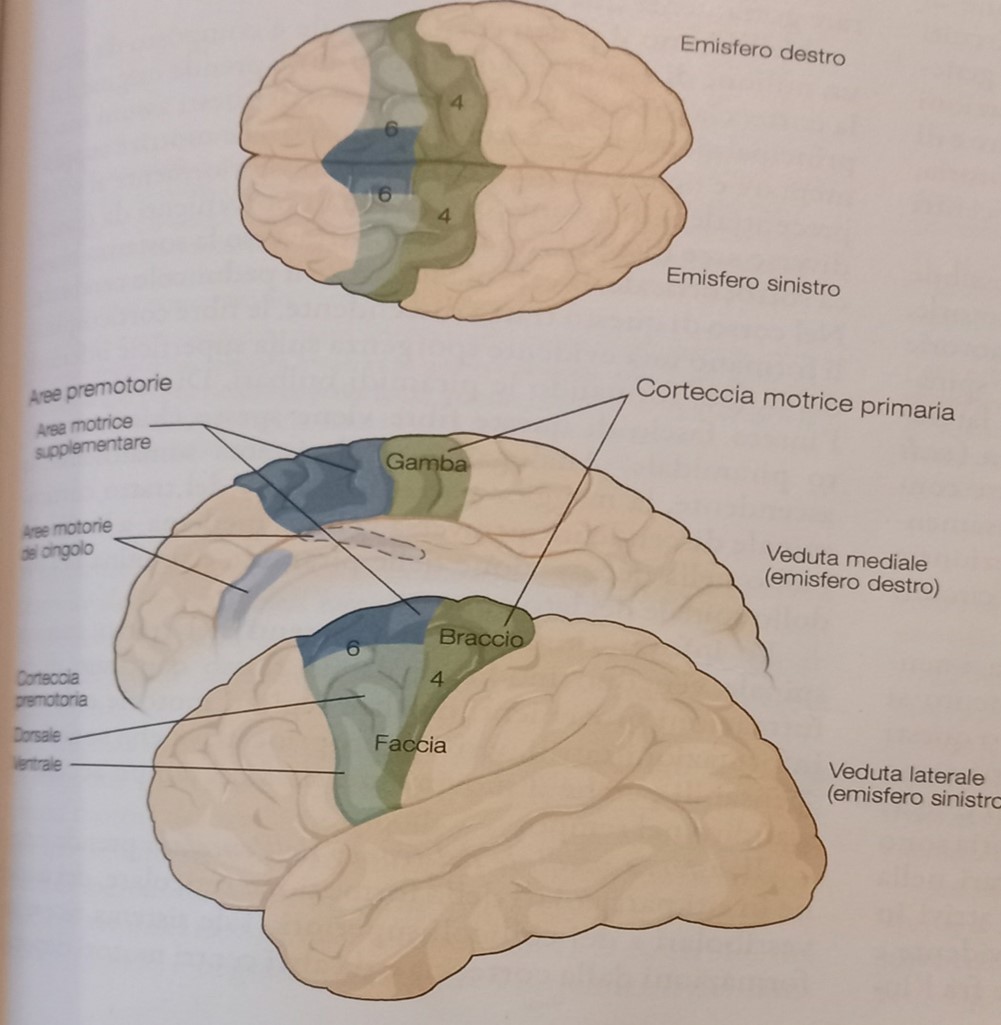
Le aree motorie identificate sono situate nel lobo frontale (pre-scissura di Rolando) e sono:
- la corteccia motrice primaria (area 4 di Brodman o M1 secondo la nuova nomenclatura)
- la corteccia premotoria (area 6 di Brodman o PM) suddivisa in
- dorsale e ventrale (PMd e PMv)
- area motrice del cingolo (superficie interna, nel solco del cingolo)
- area motrice supplementare (superficie mediale del cervello)
- la corteccia prefrontale (la zona più anteriore, quindi lo strato interno più a contatto con la fronte)
- l’area di Broca: corteccia infero-frontale
Funzionalità delle aree motorie
- corteccia motoria primaria: avvio movimenti volontari
- corteccia premotoria: supporto per i movimenti volontari, in particolar modo della bocca e della mano (area premotoria ventrale), in concomitanza di azioni finalizzate, la maggior parte dei neuroni di quest’area non si attivano quindi per i movimenti semplici.
- corteccia prefrontale: funzioni esecutive, pianificazione, emozioni, attenzione
- area di Broca: produzione del linguaggio parlato e scritto
Mappa somato-topica
In modo simile alla corteccia somato-sensitiva, in cui è stata mappata una rappresentazione isotopica delle vie sensoriali (l’homunculus somato-sensoriale), è stata individuata una mappa somato-topica motoria ma, attualmente, la concezione dell’organizzazione funzionale delle aree motrice è cambiata, in quanto nella corteccia primaria non sono rappresentate in aree motorie adiacenti semplicemente le zone periferiche adiacenti, ma i singoli muscoli e le articolazioni sono più volte rappresentate consentendo la diversificazione di movimenti semplici per azioni finalizzate anche complesse.
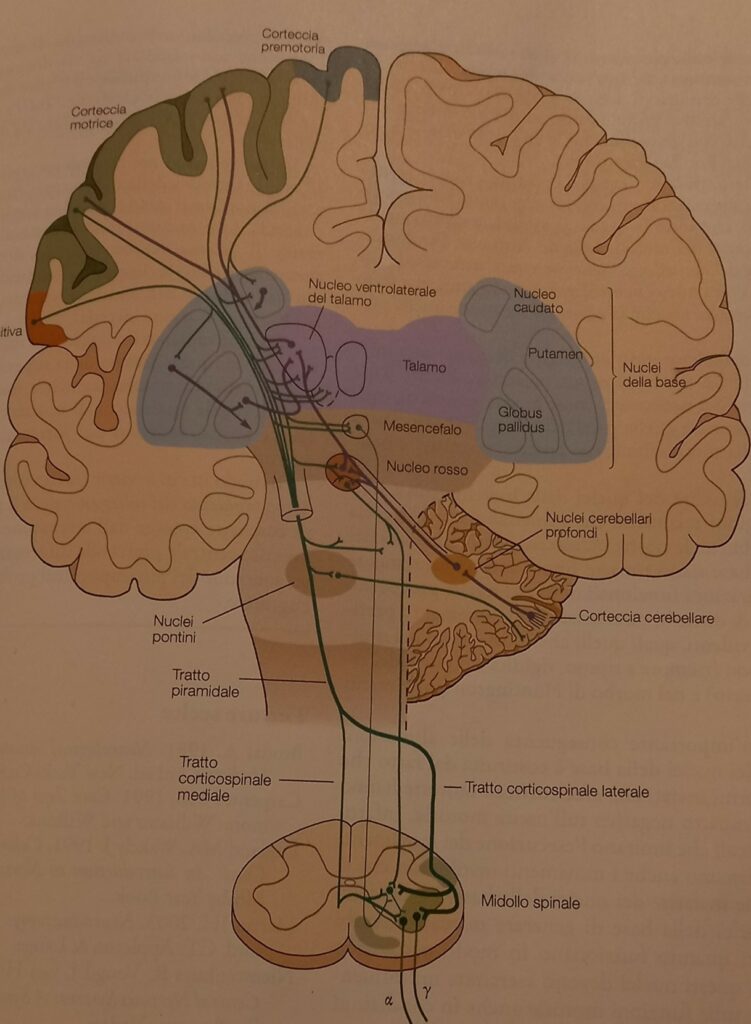
Il sistema corticospinale connette i neuroni della corteccia con motoneuroni, interneuroni e generatori di schemi motori corticali, consentendo l’integrazione e la conversione di afferenze sensitive locali in una uscita motoria.
La maggior parte delle aree motorie hanno sede nelle aree 4 e 6 di Brodman, con funzionalità diverse. L’area 4 in particolare comprende le rappresentazioni della faccia, del braccio e della gamba. Tali aree a loro volta si dividono in due aree laterali e due mediali.
Ognuna funge da stazione di raccordo ed è interconnessa alle altre per il controllo dei movimenti delle zone periferiche che rappresentano.
Nel 1930 si scoprì che era possibile evocare movimenti stimolando anche le aree pre-motorie, queste ultime risultavano prevalentemente attive in relazione a un movimento finalizzato e in assenza di un’azione motoria evidente, ossia nel periodo che intercorre tra lo stimolo comportamentale e la sua trasformazione nel movimento relativo. I neuroni con una finalità motoria risultano invece relativamente rari nella corteccia primaria. Per determinare un movimento, i segnali delle suddette aree motorie devono in ogni caso raggiungere i motoneuroni del midollo spinale.
Il tratto piramidale
Il sistema corticospinale è anche detto piramidale, in quanto le fibre nervose, nel loro decorso discendente, formano un evidente rigonfiamento di forma piramidale, a livello del margine inferiore bulbare, area di decussazione nel lato opposto del midollo spinale. Tale sistema è implicato nel controllo di tutti gli aspetti dei movimenti, ed in particolar modo nei movimenti frazionati, necessari per compiere movimenti di abilità (come scrivere o suonare il pianoforte), nello specifico è la corteccia primaria che ne consente il controllo principale, una sua lesione infatti comporta l’impossibilità di muovere indipendentemente le dita. Il tratto piramidale è pertanto deputato al controllo del movimento volontario.
In questo sistema corticospinale sono state individuate tre vie: una diretta e due indirette, mediale e laterale del tronco encefalico.
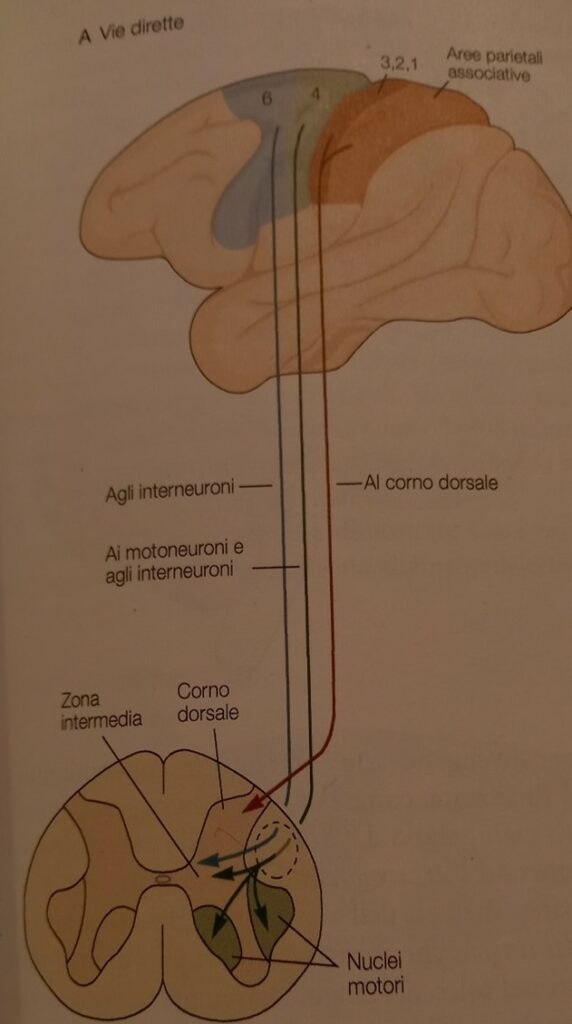
La via diretta
La via diretta prende origine dalla parte posteriore del lobo frontale e del lobo parietale (divisi dalla scissura di Rolando), il lobo frontale comprende come visto le aree 4 e 6 di Brodman, l’area 4 della corteccia motoria primaria è la sola ad avere connessioni dirette con i motoneuroni spinali, l’area 6, premotoria, ha connessione prevalentemente con gli interneuroni della zona intermedia del midollo spinale; le fibre parietali sono connesse con le corna dorsali del midollo spinale. Le informazioni motorie sono continuamente modulate da altre regioni motorie, tattili, visive e propriocettive, affinché il movimento possa essere accurato e scandito.
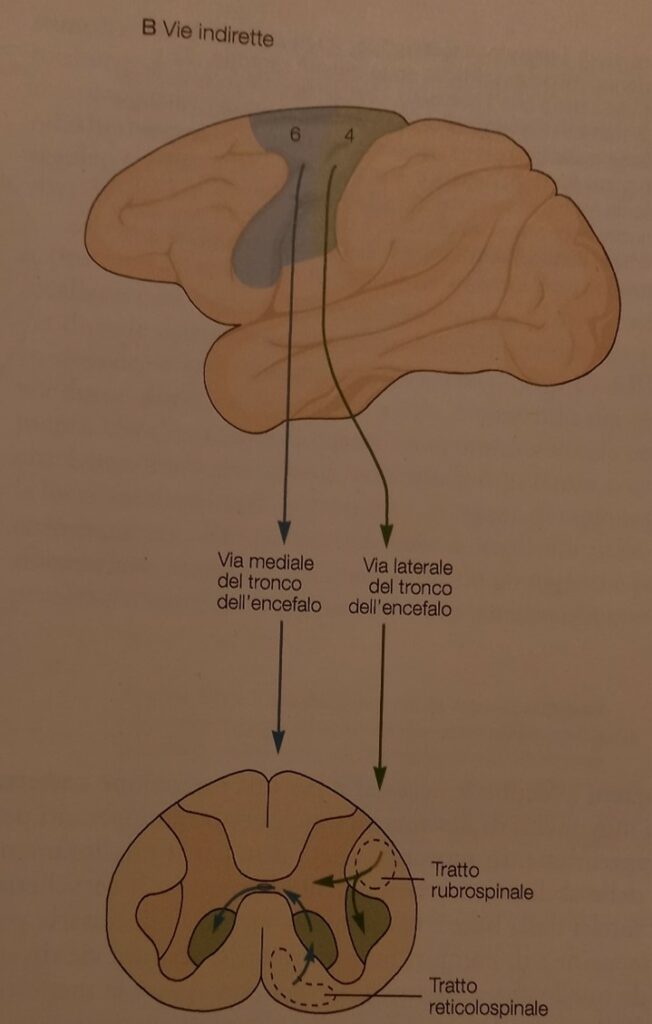
La via indiretta mediale
La via indiretta mediale del tronco encefalico collega le aree corticali con la sostanza reticolare, i nuclei vestibolari e i collicoli superiori che terminano nell’area ventromediale della sostanza grigia del midollo spinale, e prendono il nome di fasci reticolospinali, vestibolospinali e tettospinali, tale via è deputata principalmente al controllo della postura e della locomozione.
La via indiretta laterale
La via indiretta laterale è detta rubrospinale, in quanto origina dal nucleo rosso del tronco encefalico per terminare nella zona dorso-laterale del midollo spinale controlaterale.
Questa via, ricevente anch’essa afferenza dalla corteccia, è deputata al controllo dei movimenti del braccio e della mano.
Il controllo dei circuiti motori spinali
I circuiti motori spinali non vengono controllati soltanto dalle vie discendenti, i circuiti riflessi motori e i generatori di schemi motori del midollo spinale possono coordinare i movimenti stereotipati (come il cammino), senza l’intervento delle vie discendenti, ma queste li coordinano ulteriormente e possono creare nuovi schemi di attivazione dei muscoli agendo direttamente sui motoneuroni. Tale controllo corticale conferisce al movimento una flessibilità e un’accuratezza molto maggiore rispetto ai movimenti generati esclusivamente dai circuiti motori spinali.
Le aree motorie corticali e del tronco encefalico ricevono a loro volta afferenze dal cervelletto e dai nuclei della base per affinare i movimenti, sono quindi importanti per l’apprendimento motorio.
Sono inoltre collegate reciprocamente con le arre associative della corteccia parietale posteriore che costituiscono i circuiti visuo-motori. È possibile collegare azioni diverse alla vista del medesimo oggetto, alcune vie visive, infatti, sono collegate alla sola percezione, mentre altre sono implicate nella progettazione di atti motori. Le prime, le vie visive ventrali, permettono il riconoscimento degli oggetti; le seconde, le vie visive dorsali, di collocarli nello spazio e interagire con essi. Una lesione della via ventrale determina quindi la possibilità di usare correttamente un oggetto senza riconoscerlo, pertanto solo i circuiti della via dorsale sono in grado di guidare il comportamento.
La via dorsale
La via dorsale si suddivide in dorso-dorsale e ventro-dorsale.
La dorso-dorsale è implicata nel controllo dei movimenti, una lesione di questa via comporta atassia ottica, per cui il soggetto sa dove sono collocati gli oggetti ma non sa come raggiungerli.
La via ventro-dorsale, implicata nelle trasformazioni visuo-motorie, ossia per percepire gli oggetti e interagire con essi. Una lesione di questa via comporta il neglet o negligenza spaziale. In seguito ad ictus che hanno lesionato tale area il paziente non percepisce gli oggetti posti dal lato dell’emisoma colpito (a volte il lato colpito del loro stesso corpo).
Il movimento volontario
Il movimento innescato dalla scelta volontaria è una delle caratteristiche più importanti dei primati. Le azioni riflesse sono risposte più o meno stereotipate a stimoli esterni e interni,
le azioni volontarie sono le manifestazioni dell’intenzionalità a muoversi,
generate a livello corticale; per cui si può rispondere o meno a uno stimolo o rispondere in maniera diversa allo stesso, avvicinarsi alla sorgente dello stimolo, evitarlo o ignorarlo. Il cervello dell’Uomo in particolare, e dei primati in generale, è caratterizzato da una notevole espansione della corteccia cerebrale, correlata a un aumento delle capacità sensitive motorie e cognitive.
I ricercatori Sakata e Rizzolatti individuarono le aree motorie di prensione nella zona infraparietale anteriore, attraverso esperimenti condotti su scimmie addestrate a prendere oggetti diversi, registrando le attività neuronali durante lo svolgimento di tali compiti, sia alla luce sia al buio. Si evidenziarono diverse classi neuronali:
- a dominanza visiva, attivati solo durante i movimenti eseguiti alla luce
- a dominanza motoria, rispondenti durante i movimenti eseguiti sia alla luce sia al buio (quindi utilizzando criteri mnemonici)
- a dominanza visuomotoria, attivati anch’essi durante l’esecuzione di movimenti sia alla luce sia al buio, ma con scarica maggiore alla luce.
Osservarono, inoltre, che la maggior parte dei neuroni a dominanza visiva e a dominanza motoria scaricano anche in assenza di movimento di prensione ma durante l’osservazione visiva dell’oggetto, così come i neuroni che rispondono selettivamente durante la manipolazione dell’oggetto lo fanno alla sola vista dell’oggetto, in assenza di movimento. Da ciò Sakata ne dedusse che i neuroni dell’area infraparietale anteriore sono implicati nella trasformazione delle rappresentazioni sensitive degli oggetti in rappresentazioni motorie.
Successivamente, Rizzolatti e collaboratori misero in evidenza che durante i movimenti di prensione della mano si registrava l’attività di un’area della corteccia premotoria ventrale, denominata F5. Sorprendentemente, anche la maggior parte dei neuroni di quest’area, denominati canonici, si attivavano durante l’osservazione dell’oggetto anche in assenza di movimento, indipendentemente, quindi dalla preparazione alla presa dell’oggetto e in modo selettivo in base alle caratteristiche dell’oggetto (forma, dimensione e orientamento). Ciò suggerisce che la presentazione di un oggetto possa innescare la traduzione dell’oggetto in un potenziale atto motorio.
Questi studi posero le basi per l’elaborazione di un nuovo modello di elaborazione delle rappresentazioni sensitive trasformate in movimenti della mano[2]. Il modello è il seguente: lo stimolo visivo (area visiva dorsale) manda segnali all’area intraparietale anteriore che codifica la possibilità d’interazione con l’oggetto, informazione che viene inviata all’area F5 (premotoria ventrale) che codifica i possibili atti motori. Questi però non coincidono soltanto con le caratteristiche dell’oggetto ma con le opportunità pratiche di esso (affordances[3]) perché un oggetto può essere utilizzato in diversi modi, anche non convenzionali. Quindi, un modello più completo e realistico del circuito di prensione implica che le suddette aree corticali prevedano non una ma tutte le possibilità d’azione, la cui selezione sarà influenzata dall’intenzionalità del soggetto.
I neuroni a specchio
Come visto, la scarica dei neuroni canonici che avviene anche in assenza di un movimento evidente rappresenta un potenziale atto motorio, quindi una pianificazione di movimento ma senza invio del comando motorio, ciò determina la facoltà di scegliere se rispondere o meno a uno stimolo o semplicemente memorizzarlo.
All’interno dell’area F5 esiste un gruppo di neuroni denominati neuroni specchio che scaricano durante l’esecuzione di movimenti specifici (come afferrare o tenere in mano qualcosa) o quando il soggetto ne guarda un altro che esegue lo stesso movimento, ma non quando gli viene presentato un oggetto (contrariamente ai neuroni canonici), quindi è l’azione dell’altro che viene codificata come atto potenziale.
Poiché l’informazione visiva non coincide con la comprensione del significato di un’azione, probabilmente la loro funzione è farci capire l’intenzione, il significato dell’azione che stiamo osservando compiere dall’altro, sulla base della propria esperienza.
La pianificazione di un atto motorio volontario risulta essere un sistema di controllo sul movimento stesso, un’alterazione di alcune aree premotorie o motorie, infatti, possono determinare una sindrome neurologica del comportamento di utilizzazione, in conseguenza della quale il paziente afferra qualunque oggetto in modo compulsivo; o un altro disturbo neurologico che si manifesta con la difficoltà a iniziare o eseguire movimenti il cui scopo non è compreso.
In conclusione, possiamo affermare che il movimento è la risultanza delle numerose informazioni reciproche di diverse aree corticali e sottocorticali, in particolare delle aree premotorie e motoria primaria con le aree sensitive e associative corticali, che inviano, dopo averlo pianificato e avendone analizzato le varie caratteristiche con cui interagire, il comando motorio al sistema muscolo-scheletrico necessario. Ogni nuovo movimento appreso implica il coinvolgimento contemporaneo delle aree senso-motorie per aggiornare progressivamente il programma motorio, e di quella supplementare in particolare per apprenderlo attraverso la pianificazione, la genesi e il controllo delle azioni motorie sequenziali, per poi cessare definitivamente una volta che il nuovo comportamento diviene automatico, quindi appreso.
Neuroni non a specchio ed estensioni della Sigmasofia
Recenti studi hanno evidenziato che i neuroni specchio si attivano
anche in assenza di attenzione visiva
mentre un altro soggetto compie un’azione, pertanto ciò potrebbe essere spiegato avvalendosi dell’entanglement, secondo cui tutto è atomicamente e coscienzialmente legato. Poiché, secondo la Sigmasofia, la coscienza è un campo di forza esteso, indifferenziato, quindi pre-contenuto specifico (acquisito), che si avvale del cervello per essere differenziata e decodificata in contenuti acquisiti (il movimento di cui stiamo trattando nello specifico, così come le emozioni, i pensieri, le sensazioni), alcune strutture cerebrali possono attivarsi simultaneamente a distanza in più individui, anche senza collegamento diretto. Ciò potrebbe spiegare alcune capacità come la telepatia, la tele-somato-patia a livello neurale.
Lo studio del sistema motorio mette in evidenza che per l’evoluzione dell’individuo, sia filogeneticamente sia nella progressione delle tappe evolutive individuali,
l’esperienza diretta e somatica riveste un ruolo fondamentale.
Permette, infatti, di stabilire connessioni neurologiche specifiche che possono essere continuamente ampliate attraverso l’apprendimento di nuovi modelli comportamentali, grazie alle proprietà neuroplastiche del cervello stesso. Come visto, rispetto ad un oggetto, il cervello elabora tutti i potenziali atti motori che poi vengono selezionati in base alle affordances (le opportunità pratiche), quindi, sperimentare nuovi contesti, interni ed esterni, con la finalità di consapevolizzarli, elaborarli (funzione menzionata nelle precedenti lezioni in relazione allo sviluppo del sistema cognitivo-emotivo) come la Sigmasofia propone, attraverso i vari strumenti operativi da vivere, amplia notevolmente il ventaglio delle possibilità comportamentali, possibilità che una Io-psyché identificato nelle coazioni a ripetere non può assumere, appunto perché rafforza sempre gli stessi percorsi neuronali.
Approfondire la conoscenza di sé,
inoltre, necessariamente coincide con
l’intuire l’intenzionalità e riconoscere l’azione dell’altro,
come ci insegnano i neuroni specchio (che si basano sulla propria esperienza) ma, estendendo la propria consapevolezza oltre i propri confini percettivi, si può partecipare l’entanglement coscienziale, l’essere fusionali con il tutto (autonomamente fusionali) come i neuroni non a specchio sembrano confermare.
Note:
[1] Cit. Pag. 412 Principi di neuroscienze, quarta edizione italiana; E.R.Kandel, J.H.Schwartz, T.M. Jessel, S.A.Siegelbaum, A.J.Hudspeth. Casa Editrice Ambrosiana.
[2] Elaborato dai neuroscienziati Michel Arbib, Rizzolatti, Luppino.
[3] Concetto introdotto dallo psicologo statunitense James J.Gibson.
Lascia un commento